ウイルス考古学~ウイルスと哺乳類が繰り広げてきた進化的バトル~
[紹介論文] Ito J., Gifford RJ., and Sato K. Retroviruses drive the rapid evolution of mammalian APOBEC3 genes. (2020), Proc Natl Acad Sci U S A.
ゲノムに眠る「分子化石」である内在性レトロウイルスを使って、ウイルスと宿主の進化的バトルを解析した論文をご紹介いたします。
ゲノムの歴史とロマンを感じられる・・・かも!?

この記事の見出し
要約
進化の過程において、哺乳類の祖先は病原性ウイルスに対する様々な防御機構を発達させてきた。抗レトロウイルス因子の一つであるAPOBEC3タンパク質は、guanine-to-adenine (G-to-A) 変異をウイルスゲノムに導入することで、様々なレトロウイルスの複製を強力に抑制する。興味深いことに、APOBEC3ファミリー遺伝子は哺乳類の進化過程において遺伝子重複を繰り返し、急速に進化してきたことが知られている。このようなAPOBEC3ファミリー遺伝子の急速な進化は、レトロウイルスによる侵略に対抗するために引き起こされたと推測されてきたものの、その具体的な証左は示されてこなかった。一方で、哺乳類のゲノム中には、過去のレトロウイルス感染の痕跡である内在性レトロウイルス (endogenous retrovirus; ERV) が大量に存在する。ERVは古代のレトロウイルスの「分子化石」であるため、ゲノム中に残るERV配列を解析することで、宿主の祖先–ウイルス間の進化的攻防を解明するための手掛かりを得ることができる。
本研究では、160種類の哺乳類を対象とした比較ゲノム解析を行うことで、レトロウイルスとAPOBEC3ファミリー遺伝子の間に起きた進化的攻防を明らかにすることを試みた。各哺乳類のゲノムにおいてAPOBEC3ファミリー遺伝子およびERV配列を網羅的に同定・計数し、さらにはERV配列中に蓄積したG-to-A変異を定量した。その結果、ゲノム中にERVを多く含み、過去に多くのレトロウイルス感染を経験したと考えられる動物種ほど、より多種多様なAPOBEC3ファミリー遺伝子を持つことが明らかとなった。また、多数のAPOBEC3ファミリー遺伝子を持つ動物種ほど、ERV配列中により多くのG-to-A変異を蓄積していることが明らかとなった。さらに、霊長類において、大量のレトロウイルス・ERVがゲノムを侵略した時期に、APOBEC3ファミリー遺伝子の増幅が起こったことが明らかとなった。
以上の結果から、レトロウイルスの侵略が哺乳類におけるAPOBEC3ファミリー遺伝子の増幅のきっかけとなったこと、およびAPOBEC3ファミリー遺伝子の増加がレトロウイルスへの攻撃の強化に繋がったことが示唆された。本研究は、ウイルスと宿主の長期間にわたる進化的攻防を理解する上での貴重なモデルケースを提供するものである。
導入
内在性レトロウイルス(endogenous retrovirus; ERV)は、古代のレトロウイルスが宿主の生殖細胞に感染し、宿主ゲノムに組み込まれることで生じた「ウイルス感染・侵略の痕跡」である1。現存する哺乳類において、ERV由来の配列はゲノムの大きな割合(~12%)を占めている。このことは、進化の過程において哺乳類の祖先が大量のレトロウイルス感染に暴露されてきたことを示唆している。また、いったん宿主生殖細胞への侵入を果たし、ERVへと進化したレトロウイルスは、トランスポゾンの一種として増殖を繰り返し、宿主のゲノムをさらに侵略していく。このようなレトロウイルス・ERVの感染・侵略に対抗するため、哺乳類の祖先はさまざまなウイルス感染防御機構を進化させてきた。
レトロウイルスの複製を阻害する遺伝子のひとつとして、APOBEC3ファミリー遺伝子が知られている2。APOBEC3タンパク質は、シチジン脱アミノ化酵素活性を持っており、レトロウイルスの相補鎖DNA(complementary DNA; cDNA; アンチセンス鎖)にcytosine-to-uracil変異を導入する。その結果、ウイルスゲノム(センス鎖)にguanine-to-adenine (G-to-A) 変異が導入され、レトロウイルスの複製を強力に抑制する。APOBEC3ファミリー遺伝子は、ヒト免疫不全ウイルス (HIV-1) 等の“外来性”のレトロウイルスに対する防御遺伝子として有名だが、ERVの増殖に対しても抑制的に働きうることが報告されている3。
興味深いことに、APOBEC3ファミリー遺伝子の数は、哺乳類の種間において大きく異なることが知られている4。例として、ヒトは7つのAPOBEC3ファミリー遺伝子を持つものの、マウスは1つしか持っておらず、有袋類に至っては1つも持っていない。APOBEC3ファミリー遺伝子の進化にまつわる魅力的な仮説のひとつに、「APOBEC3ファミリー遺伝子のコピー数の増加は、レトロウイルスまたはERVの侵略に対抗するために起こった」というものがある。しかしこの仮説を支持する具体的な証拠は未だ提示されていない。
ゲノム中に存在するERV配列は、過去に存在したウイルスの「分子化石」のようなものである。そのため、ERV配列を調べることで、1) どのような種類のウイルスが、2) どの生物に、3) いつ、 4) どの程度感染していたかを知ることができる。さらには、ERV配列を精査することで、長期にわたるウイルスと宿主の進化的攻防を解明するための手がかりを得ることができるかもしれない。特にAPOBEC3ファミリー遺伝子は、攻撃したウイルスの配列中に分子的な痕跡(G-to-A変異)を残すという稀有な性質があるため、ERV配列におけるG-to-A変異の蓄積量を調べることで、ERVあるいはレトロウイルスが過去にどの程度APOBEC3から攻撃を受けたか推定することができる。そこで本研究では、160種類の哺乳類を対象とした大規模な比較ゲノム解析を行い、1) APOBEC3を含む「AID/APOBECファミリー遺伝子*」の網羅的同定、2) ERV配列の網羅的同定、および3) ERV配列に蓄積したG-to-A変異の定量を行った。そして、哺乳類の進化過程におけるAPOBEC3ファミリー遺伝子とレトロウイルスの進化的軍拡競争の描出を試みた。
*AID/APOBECファミリー遺伝子
シチジン脱アミノ化酵素活性を持つ。AID, APOBEC1, APOBEC2, APOBEC3, およびAPOBEC4が含まれる。抗体の成熟(AID)、脂質代謝(APOBEC1)、およびウイルス防御(APOBEC3)等、様々な生理機能に関わる。
結果および考察
1, 哺乳類におけるAID/APOBECファミリー遺伝子の網羅的同定
160種類の哺乳類のゲノム配列を対象に配列相同性(tBLASTn)検索を行い、ゲノム中に存在するAID/APOBECファミリー遺伝子に類似する配列を網羅的に同定した。その結果、1,420個のAID/APOBECファミリー遺伝子が同定された。各動物種において同定されたAID/APOBECファミリー遺伝子の数を図1に示す。APOBEC3以外のAID/APOBECファミリー遺伝子は、各動物種において概ね1コピーずつしか存在しなかった。一方で、APOBEC3ファミリー遺伝子(A3/Z1、A3/Z2、およびA3/Z3に細分化される)は、霊長目、コウモリ目、ウマ目、ゾウ目等において顕著にコピー数が増加していた。以上のように、哺乳類の複数の系統においてAOBEC3ファミリー遺伝子の増加が起こったことが明らかとなった。

図1 哺乳類の各種におけるAID/APOBECファミリー遺伝子のコピー数。
2, ERV配列の網羅的同定およびERVにおけるG-to-A変異蓄積量の定量
配列相同性検索(RepeatMakser)により、ゲノム中に存在するERV配列を網羅的に同定したところ、ERVの挿入量は哺乳類の系統間において大きく異なっていることが明らかとなった(図2A)。この結果は、過去に受けたレトロウイルスまたはERVの侵略の度合が、哺乳類の系統間において大きく異なっていることを示唆している。
レトロウイルスおよびERVが、APOBEC3から過去に受けた攻撃の程度を推定するために、ERV配列中に蓄積したG-to-A変異の量を定量した。APOBEC3ファミリー遺伝子は、レトロウイルスのセンス鎖特異的にG-to-A変異を導入するという性質を持つ。そこで本研究では、ERV配列のセンス鎖・アンチセンス鎖間におけるG-to-A変異速度の比を計算し、これをAPOBEC3によるG-to-A変異蓄積スコアと定義した。図2Bに示す通り、ゲノム中のERV配列におけるG-to-A変異蓄積量は、哺乳類の系統間において大きく異なっていた。この結果は、APOBEC3によるERVへの攻撃の程度が哺乳類間において異なっていたことを示唆している。
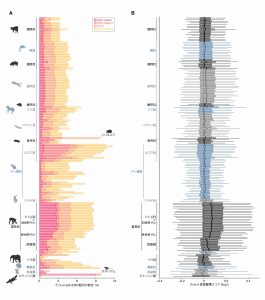
図2 ゲノム中のERV挿入量(A)およびERV配列におけるG-to-A変異の蓄積量(B)。
3, レトロウイルス・ERVの侵略とAPOBEC3ファミリー遺伝子の増幅の関連
哺乳類各種のゲノムに組み込まれたERVの量と、APOBEC3ファミリー遺伝子の数を比較したところ、正の相関が見られた(図3A)。すなわち、ゲノム中にERVを多く含み、過去に多くのレトロウイルス・ERVの侵略を経験したと考えられる動物種ほど、より多くのAPOBEC3ファミリー遺伝子を持つことが明らかとなった。この結果は、レトロウイルス・ERVの侵略がAPOBEC3ファミリー遺伝子の遺伝子数増加の一因となったことを示唆している。
哺乳類の各種が持つAPOBEC3ファミリー遺伝子の数と、ERV配列中に蓄積したG-to-A変異の量の関連を解析したところ、正の相関が見られた(図3B)。すなわち、APOBEC3ファミリー遺伝子を多く持つ動物種ほど、ERV配列中に多くのG-to-A変異を蓄積していることが明らかとなった。この結果は、APOBEC3ファミリー遺伝子の増加がERVへの攻撃の強化に繋がったことを示唆している。
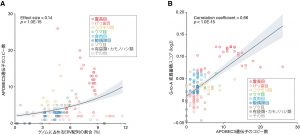
図3 ゲノム中のERV挿入量、APOBEC3ファミリー遺伝子のコピー数、およびERV配列におけるG-to-A変異の蓄積量の関連解析。(A)ERV挿入量とAPOBEC3ファミリー遺伝子のコピー数の関連。(B)APOBEC3ファミリー遺伝子のコピー数とG-to-A変異の蓄積量の関連。
レトロウイルスの侵略とAPOBEC3ファミリー遺伝子数増加の関連性をより明らかにするために、両者の起こった進化的な時期を比較した。ここでは特に、レトロウイルス・ERVの侵略とAPOBEC3ファミリー遺伝子数の増加が顕著に見られた霊長類に注目し解析を行った(図4A)。その結果、ERVのゲノムへの挿入は、真猿類(ヒト上科、旧世界ザル、および新世界ザルを含む)の共通祖先(約5,000万年前)において活発に起こったことが明らかとなった。一方で、ERVの挿入は、同時期の原猿類(メガネザル、キツネザル、ブッシュベイビー等を含む)の祖先にはほとんど起こっていなかった。また、真猿類は多数のAPOBEC3ファミリー遺伝子を持つ一方、ほとんどの原猿類はごく少数のAPOBEC3ファミリー遺伝子を持っていることが分かった(図4B)。このことは、霊長類におけるAPOBEC3ファミリー遺伝子の顕著な増加は、真猿類・原猿類の分岐後に真猿類の共通祖先において起こったことを示唆している。すなわち、霊長類において、レトロウイルス・ERVの顕著な侵略が見られたのと同時期・同系統において、APOBEC3ファミリー遺伝子の増幅が起こったことが示された。以上の結果は、レトロウイルス・ERVの侵略がAPOBEC3ファミリー遺伝子の遺伝子数増加の一因となったことを示唆している。

図4 霊長類におけるレトロウイルスの侵略時期とAPOBEC3ファミリー遺伝子増幅の時期の関連。(A)各年代・各種におけるERVの推定挿入量を表すヒートマップ。(B)霊長類各種におけるAPOBEC3ファミリー遺伝子のコピー数。
総括
本研究結果から、レトロウイルス・ERVの侵略がAPOBEC3ファミリー遺伝子増加の一因となったことを示唆する結果が得られた。また、APOBEC3ファミリー遺伝子の増加が、レトロウイルスに対する攻撃の強化に繋がったことを示唆する結果が得られた。レトロウイルスがAPOBEC3ファミリー遺伝子の進化を駆動してきたという仮説、あるいはウイルスが抗ウイルス遺伝子の進化を駆動してきたという仮説は、これまで活発に議論されてきたものの、具体的証拠に乏しかった。特に、数千万年のタイムスケールにおけるウイルスと抗ウイルス遺伝子の進化攻防については、これまで解析することは難しかった。本研究では、「分子化石」であるERV、およびその化石に傷(G-to-A変異)を残す性質を持つAPOBEC3ファミリー遺伝子に着目することで、ウイルスと宿主の長期間にわたる進化的攻防の歴史を再構築することに成功した。むろん、ERVは不完全な化石記録であるに過ぎず、現在ゲノム中に観測されるERVは、過去に感染した膨大なレトロウイルスのごく一部が標本抽出されたものに過ぎない。それでもなお、本研究の示すレトロウイルスとAPOBEC3ファミリー遺伝子の進化的攻防のモデルケースは、今後ウイルスと抗ウイルス遺伝子の進化的攻防モデルを議論する上での貴重な論拠の一つとなるだろう。
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
研究を始めたきっかけ
本研究を始めたきっかけは若干入り組んでいます。もともとのきっかけは、佐藤佳先生が2015年の冬に共著者のRobert Gifford先生(グラスゴー大学)の研究室へ短期留学したことでした。その際に佐藤先生とGifford先生は、「哺乳類におけるAPOBEC3ファミリー遺伝子を網羅的に同定する」という仕事をされましたが、諸事情によりこの仕事はお蔵入りとなっていました。2018年の秋、佐藤先生と著者(伊東)が再びグラスゴー大学を訪れた際、Gifford先生が古いファイルを持ち出してきて、「APOBEC3ファミリー遺伝子の仕事をまとめないか」と提案されました。その後、大学や酒場(図5)でのなんやかんやの議論の末、「APOBEC3ファミリー遺伝子、ERVの量、G-to-A変異の関連を見るんや!」ということに落ち着き、この仕事がスタートしました。ちなみにGifford先生は、ERV配列に基づくウイルス進化解析の草分け的な仕事をされてきた先生です。

図5 グラスゴーの酒場にて。左から佐藤佳先生、Robert Gifford先生、著者(伊東)。著者は写真を撮ると大体目を瞑っている。
苦労した点
先生方のサポートもあり、この仕事は半年程度で上手くまとまってくれたので、苦労した点は正直あまりありません(他の仕事もこの調子で行けばいいのですが…)。強いていうなら、1) G-to-A変異の蓄積量を評価する方法(G-to-A変異蓄積スコア)を考える点と、2) 仕事としてのまとめ方です。2) に関しては、2019年5月開催の国際学会にて発表するという明確なタイムリミットがあったことがよかったように思われます。
参考文献
1 Feschotte, C. & Gilbert, C. Endogenous viruses: insights into viral evolution and impact on host biology. Nat Rev Genet 13, 283-296, doi:10.1038/nrg3199 (2012).
2 Harris, R. S. & Dudley, J. P. APOBECs and virus restriction. Virology 479-480, 131-145, doi:10.1016/j.virol.2015.03.012 (2015).
3 Treger, R. S. et al. Human APOBEC3G Prevents Emergence of Infectious Endogenous Retrovirus in Mice. J Virol 93, doi:10.1128/jvi.00728-19 (2019).
4 Nakano, Y. et al. A conflict of interest: the evolutionary arms race between mammalian APOBEC3 and lentiviral Vif. Retrovirology 14, 31, doi:10.1186/s12977-017-0355-4 (2017).
謝辞
本研究はRobert J. Gifford博士(グラスゴー大学)と共同で行われました。
また、本研究は菅波麻衣氏(東京大学)の技術協力のもと行われました。
おまけ・研究室紹介:
東京大学 医科学研究所「システムウイルス学研究室」について
「システムウイルス学」とは、著者(伊東)の上司である佐藤佳先生(図6)とその恩師である京都大学の小柳義夫先生が作られた言葉で、「分野横断的な技術・知識を駆使し、ウイルスをシステム(総体)として理解することを目指す学問」という意味だそうです。著者個人としては、「いろいろなウイルスを様々なアプローチから研究する学問」と平たく捉えています。実際、ラボメンバー(図7)の扱うウイルスは様々です。研究室のメインテーマは、ヒトにAIDSを引き起こす「ヒト免疫不全ウイルス-1(HIV-1)」ですが、今回ご紹介させて頂いたような内在性レトロウイルスの研究をしている人もいますし、ウイルスの種類に拘らず、サンプル中に含まれるウイルスを包括的に解析する手法「Virome」を用いて研究を行っている人もいます。また、研究に用いているアプローチも多種多様です。細胞生物学的手法(Wet解析)を専門とするメンバーもいれば、バイオインフォマティクス(Dry解析)を得意とするメンバーもいますし、どちらの技術を持つ人もいます。医学的な興味が強い人も、分子生物学あるいは進化学的な観点から研究を行なっている人もいます。様々な興味・技術を持つメンバーが互いに協力し合い、日々研究を進めています。また研究室を主催する佐藤先生は、広い興味・視野を持ち、フットワークが軽く、とても面倒見の良い先生です。大学院生・ポスドクも募集中ですので、興味のある方はぜひ研究室に遊びに来てください。

図6 佐藤佳先生。新学術領域研究「ネオウイルス学」(http://neo-virology.org/)にて凄腕カメラマンに撮影してもらった。

図7 システムウイルス学研究室のメンバー。写真は佐藤先生のお誕生日会。
研究室HP:
http://www.ims.u-tokyo.ac.jp/SystemsVirology
研究室Twitter & Facebook:
@SystemsVirology
佐藤先生 メールアドレス:
ksato@ims.u-tokyo.ac.jp
著者情報:
伊東潤平(イトウジュンペイ)
東京大学医科学研究所 感染症国際研究センター システムウイルス学分野
学振特別研究員(PD)
ゲノム生物学、生物情報学、ウイルス学が専門で、内在性レトロウイルスが大好きです。内在性レトロウイルスの駆動する生物進化と遺伝子制御ネットワークについて研究しています。
佐藤 佳(サトウ ケイ)
東京大学医科学研究所 感染症国際研究センター システムウイルス学分野
准教授(研究主宰者)











